РНК-полимераза
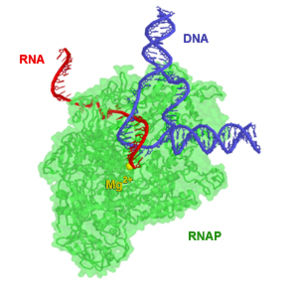

РНК-полимераза — фермент, осуществляющий синтез молекул РНК. В узком смысле, РНК-полимеразой обычно называют ДНК-зависимые РНК-полимеразы, осуществляющие синтез молекул РНК на матрице ДНК, то есть осуществляющие транскрипцию. Ферменты класса РНК-полимераз очень важны для функционирования клетки, поэтому они имеются во всех организмах и во многих вирусах. Химически РНК-полимеразы являются нуклеотидил-трансферазами, полимеризующими рибонуклеотиды на 3′-конце цепи РНК.
Содержание
История изучения
Нобелевская премия по химии в 2006 году была присуждена Роджеру Корнбергу за получение точных изображений молекул РНК-полимеразы в различные моменты процесса транскрипции. [3]
Управление транскрипцией
![]()
Управление процессом транскрипции генов позволяет контролировать экспрессию генов и таким образом позволяет клетке адаптироваться к изменяющимся условиям внешней среды, поддерживать метаболические процессы на должном уровне, а также выполнять специфические функции, необходимые для существования организма. Неудивительно, что действие РНК-полимеразы очень сложно и зависит от множества факторов (так, у Escherichia coli идентифицировано более 100 факторов, тем или иным способом влияющих на РНК-полимеразу [4] ).
РНК-полимераза начинает транскрипцию с особых участков ДНК, называемых промоторами и производит цепочку РНК, комплементарную соответствующей части нити ДНК.
Процесс наращивания молекулы РНК нуклеотидами называется элонгацией. В эукариотических клетках РНК-полимераза может собирать цепочки из более 2,4 млн элементов (например, такую длину имеет полный ген белка дистрофина).
РНК-полимераза завершает формирование цепочки РНК, когда встречает в ДНК специфическую последовательность, называемую терминатором.
РНК-полимераза производит следующие разновидности РНК:
РНК-полимераза осуществляет синтез с нуля. Это возможно вследствие того, что взаимодействие начального нуклеотида гена и РНК-полимеразы позволяет ей закрепиться на цепочке и обрабатывать следующие нуклеотиды. Это отчасти объясняет, почему РНК-полимераза обычно начинает транксрипцию с АТФ, за которым следует ГТФ, УТФ и затем ЦТФ. В отличие от ДНК-полимеразы РНК-полимераза обладает также геликазным действием.
Действие РНК-полимеразы
Связывание и инициирование транскрипции

После связывания с ДНК структура РНК-полимеразы превращается из закрытой в открытую. Это превращение включает в себя разделение моноспиралей ДНК с образованием раскрученного участка длиной около 13 шагов. Рибонуклеотиды затем собираются в цепочку в соответствии с базовой нитью ДНК, используемой в качестве шаблона. Суперскрученность молекул ДНК играет существенную роль в деятельности РНК-полимеразы: поскольку участок ДНК перед РНК-полимеразой раскручен, в нем существуют положительные компенсационные супервитки. Участки ДНК позади РНК-полимеразы снова закручиваются и в них присутствуют отрицательные супервитки.
Элонгация
Во время элонгационной фазы транскрипции происходит добавление рибонуклеотидов к цепи и переход от структуры РНК-полимеразного комплекса от открытой к транскрипционной. По мере сборки молекулы РНК участок ДНК перед РНК-полимеразой раскручивается далее, и 13-парный открытый комплекс превращается в 17-парный транскрипционный комплекс. В этот момент промотор (участок ДНК −10…-35 шагов) завершается, и σ-фактор отделяется от РНК-полимеразы. Это позволяет остальному РНК-полимеразному комплексу начать движение вперед, так как σ-фактор удерживал его на месте.
17-парный транскрипционный комплекс содержит гибрид ДНК и РНК, содержащий 8 пар оснований — 8-шаговый участок РНК, соединенный с шаблонной цепью ДНК. По мере выполнения транскрипции рибонуклеотиды добавляются к 3′-концу собираемой РНК, и РНК-полимеразный комплекс движется по цепи ДНК. Хотя в РНК-полимеразе не обнаружено свойств, характерных для 3′-экзонуклеазы, аналогичных проверочной деятельности ДНК-полимеразы, есть свидетельства того, что РНК-полимераза останавливается и корректирует ошибки в случаях ошибочного формирования пар оснований ДНК-РНК.
Терминация
Терминация транскрипции РНК может быть ρ-независимой либо ρ-зависимой.
ρ-независимая терминация осуществляется без помощи ρ-фактора. Транскрипция палиндромного участка ДНК приводит к формированию шпильки из РНК, зацикленной и связанной с самой собой. Эта шпилька богата гуанином и цитозином, что делает ее более стабильной, нежели гибрид ДНК-РНК. В результате 8-парный гибрид ДНК-РНК в транскрипционном комплексе сокращается до 4-парного. В случае если эти 4 последние пары оснований составлены слабыми аденином и уридином, молекула РНК отделяется. [5]
Бактериальная РНК-полимераза
У бактерий один и тот же фермент катализирует синтез трёх типов РНК: мРНК, рРНК и тРНК.
РНК-полимераза — достаточно большая молекула. Основной фермент содержит 5 субъединиц (
Для связывания с промоторными областями ДНК, основной фермент нуждается в еще одной субъединице — сигма (σ). Сигма-фактор значительно снижает сродство РНК-полимеразы к неспецифичным областям ДНК, и в то же время повышает ее чувствительность к определенным промоторам, в зависимости от своей структуры. С его помощью транскрипция начинается с нужного участка ДНК.
Полный голоэнзим таким образом состоит из 6 субъединиц: α2ββ’σω (
480 кДа). В структуре РНК-полимеразы присутствует канавка длиной 55 Å (5,5 нм) и шириной 25 Å (2,5 нм). Именно в эту канавку помещается двойная спираль ДНК, имеющая ширину 20 Å (2 нм). На длине канавки укладывается 16 нуклеотидов.
Молекулы РНК-полимеразы не растворены в цитоплазме. Когда РНК-полимераза не используется, она связывается с неспецифичными областями ДНК в ожидании открытия активного промотора.
Транскрипционные кофакторы
Существуют белки, связывающиеся с РНК-полимеразой и влияющие на ее поведение. Например greA и greB из E. coli усиливают способность РНК-полимеразы расщеплять шаблон РНК у растущего конца цепи. Такое расщепление может «спасти» застрявшую молекулу РНК-полимеразы, а также, вероятно, участвует в устранении ошибок сборки цепи РНК.
Отдельный кофактор Mfd задействован в транскрипционном восстановлении ДНК. Во время этого процесса РНК-полимераза обнаруживает поврежденные участки ДНК и привлекает другие ферменты для ее восстановления.
Многие другие кофакторы обладают регулирующим влиянием, заставляя РНК-полимеразу экспрессировать или не экспрессировать определенные гены.
РНК-полимераза в эукариотических клетках
Эукариоты обладают различными типами РНК-полимераз, классифицируемыми по типам РНК, которые они производят:
Существуют также и другие типы РНК-полимеразы, используемые в митохондриях и хлоропластах. Молекулярная масса этих ферментов составляет величину порядка 500 000. Они различаются по чувствительности к альфа-аманитину. РНК-полимераза I нечувствительна к нему, РНК-полимераза III умеренно чувствительна, а РНК-полимераза II сильно ингибируется им. [9]
РНК-полимераза у архей
Археи используют один вид РНК-полимеразы, который тем не менее очень похож на три основных типа РНК-полимераз у эукариот. Некоторые ученые предполагают, что архейная РНК-полимераза в определенном приближении может являться эволюционным предком специализированных эукариотических полимераз. [10]
РНК-полимераза у вирусов
Многие вирусы содержат РНК-полимеразу. Пожалуй, наиболее хорошо изученная вирусная РНК-полимераза имеется у бактериофага Т7. Эта РНК-полимераза, состоящая из одной субъединицы, похожа на митохондриальную и хлоропластную, а также на ДНК-полимеразу. [11] Считается, что большинство вирусных полимераз произошли от ДНК-полимеразы, а не от сложных многокомпонентных РНК-полимераз.
Вирусные полимеразы очень многочисленны. Многие из них могут использовать в качестве шаблона РНК, а не ДНК, как, например, у вирусов с двуцепочечной РНК или с одноцепочечной РНК негативной полярности. Некоторые вирусы с одноцепочечной РНК позитивной полярности также содержат РНК-зависимые РНК-полимеразы. [12]
Функциональные области
C-концевой домен РНК-полимеразы
Инициирование транскрипции
5′-кэпирование
С-концевой домен также является местом связывания комплекса кэпирования. У эукариот после синтеза 5′-конца мРНК фосфатаза концевой фосфат с 5′-конца полирибонуклеотида фермент гуанозинтрансфераза присоединяет к нему гуанозинмонофосфат. При этом образуется 5′,5′-трифосфатная связь. Кэпирующий комплекс затем диссоциирует от мРНК, 5′-кэп из ГТФ связывается с кэп-связывающим комплексом, C-концевого домена РНК-полимеразы. 5′-кэп в структуре мРНК эукариот имеет большое значение для связывания молекул мРНК с рибосомами, а также предотвращает деградацию РНК.
Сплайсосома
Углекисло-концевой домен РНК-полимеразы также является областью связывания со сплайсосомными факторами, участвующими в процессе сплайсинга РНК. Эти факторы способствуют осуществлению сплайсинга и удаление интронов в процессе транскрипции РНК.
Мутация в C-концевом домене
Был проведен ряд исследований поведения РНК-полимеразы при удалении определенных аминокислот из ее C-концевого домена. Показано, что мутации усечения C-концевого домена РНК-полимеразы II влияют на ее способность начинать транскрипцию набора генов in vivo, снижая чувствительность к активационным последовательностям этих генов.
Очистка РНК-полимеразы
РНК-полимераза может быть выделена следующими способами:
А также комбинациями вышеуказанных методов.
| Поиск | |
|---|---|
| ЧВК | статьи |
| PubMed | статьи |
| NCBI | белки |

В молекулярная биология, РНК-полимераза (сокращенно RNAP или РНКапол, и официально ДНК-направленная РНК-полимераза), является фермент который синтезирует РНК из ДНК шаблон.
Использование фермента геликаза, RNAP локально открывает двухцепочечную ДНК, так что одна цепь экспонированного нуклеотиды может использоваться в качестве шаблона для синтеза РНК, процесса, называемого транскрипция. А фактор транскрипции и связанная с ней транскрипция посреднический комплекс должен быть прикреплен к Сайт связывания ДНК называется промоутер регион до того, как RNAP сможет инициировать раскручивание ДНК в этом положении. РНКП не только инициирует транскрипцию РНК, но и направляет нуклеотиды в нужное положение, облегчает прикрепление и удлинение, имеет встроенные возможности корректуры и замены, а также возможность распознавания прерывания. В эукариоты, RNAP может строить цепи длиной до 2,4 миллиона нуклеотидов.
RNAP производит РНК, которая функционально предназначена либо для белка кодирование, т.е. информационная РНК (мРНК); или некодирование (так называемые «гены РНК»). Существует по крайней мере четыре функциональных типа генов РНК:
РНК-полимераза необходима для жизни и встречается у всех живых существ. организмы и много вирусы. В зависимости от организма РНК-полимераза может быть белковый комплекс (многосубъединичная RNAP) или состоит только из одной субъединицы (односубъединичная RNAP, ssRNAP), каждая из которых представляет независимую линию. Первый находится в бактерии, археи, и эукариоты одинаково, имея схожую основную структуру и механизм. [1] Последний находится в фаги а также эукариотические хлоропласты и митохондрии, и относится к современным ДНК-полимеразы. [2] Эукариотические и архейные РНКП имеют больше субъединиц, чем бактериальные, и контролируются по-разному.
Бактерии и археи имеют только одну РНК-полимеразу. У эукариот есть несколько типов ядерных РНКП, каждый из которых отвечает за синтез определенного подмножества РНК:
Содержание
Структура


2006 год Нобелевская премия по химии был присужден Роджер Д. Корнберг для создания подробных молекулярных изображений РНК-полимеразы на различных этапах процесса транскрипции. [3]
В большинстве прокариоты, один вид РНК-полимеразы транскрибирует все типы РНК. «Ядро» РНК-полимеразы из Кишечная палочка состоит из пяти субъединиц: две альфа (α) субъединицы 36кДа, субъединица бета (β) размером 150 кДа, первичная субъединица бета (β ‘) размером 155 кДа и малая субъединица омега (ω). Фактор сигма (σ) связывается с ядром, образуя холофермент. После начала транскрипции фактор может развязаться и позволить ферменту продолжить свою работу. [4] [5] Комплекс ядерной РНК-полимеразы образует структуру «клешня краба» или «зажим-челюсть» с внутренним каналом, проходящим по всей длине. [6] РНК-полимеразы эукариот и архей имеют схожую структуру ядра и работают аналогичным образом, хотя имеют много дополнительных субъединиц. [7]
Все RNAP содержат металл кофакторы, особенно цинк и магний катионы, которые помогают в процессе транскрипции. [8] [9]
Функция
![]()
Контроль за процессом ген транскрипция влияет на паттерны экспрессия гена и, таким образом, позволяет ячейка адаптироваться к изменяющейся среде, выполнять особые роли в организме и поддерживать основные метаболические процессы, необходимые для выживания. Поэтому неудивительно, что активность РНКП длительная, сложная и строго регулируется. В кишечная палочка бактерии, более 100 факторы транскрипции были идентифицированы, которые модифицируют активность RNAP. [10]
RNAP может инициировать транскрипцию в определенных последовательностях ДНК, известных как промоутеры. Затем он производит цепь РНК, которая дополнительный к цепи ДНК-матрицы. Процесс добавления нуклеотиды к цепи РНК называется удлинением; у эукариот RNAP может строить цепочки длиной до 2,4 миллиона нуклеотиды (полная длина дистрофин ген). RNAP будет преимущественно высвобождать свой транскрипт РНК в определенных последовательностях ДНК, кодируемых на концах генов, которые известны как терминаторы.
Продукция RNAP включает:
RNAP выполняет de novo синтез. Это возможно благодаря специфическим взаимодействиям с инициирующим нуклеотидом, которые жестко удерживают RNAP на месте, облегчая химическую атаку на поступающий нуклеотид. Такие специфические взаимодействия объясняют, почему RNAP предпочитает запускать транскрипты с АТФ (за которым следуют GTP, UTP и затем CTP). В отличие от ДНК-полимераза, RNAP включает геликаза активности, поэтому для раскрутки ДНК не требуется отдельный фермент.
Действие
Инициация
После связывания с ДНК РНК-полимераза переключается с закрытого комплекса на открытый. Это изменение включает разделение цепей ДНК с образованием развернутого участка ДНК размером примерно 13 п.н., называемого «пузырь транскрипции». Суперспирализация играет важную роль в активности полимеразы из-за раскручивания и перемотки ДНК. Поскольку участки ДНК перед RNAP разматываются, возникают компенсаторные положительные суперспирали. Области за RNAP перемотаны, и присутствуют отрицательные суперспирали. [13]
Побег промоутера
Затем РНК-полимераза начинает синтезировать исходный гетеродуплекс ДНК-РНК с рибонуклеотидными основаниями, спаренными с цепью матричной ДНК в соответствии с взаимодействиями пар оснований Уотсона-Крика. Как отмечалось выше, РНК-полимераза контактирует с промоторной областью. Однако эти стабилизирующие контакты подавляют способность фермента получать доступ к ДНК дальше по течению и, таким образом, синтез полноразмерного продукта. Чтобы продолжить синтез РНК, РНК-полимераза должна ускользнуть от промотора. Он должен поддерживать контакты промотора, раскручивая более низкую ДНК для синтеза, «хруст» ДНК в инициирующий комплекс. [14] Во время перехода от промотора РНК-полимераза считается «промежуточным продуктом стресса». Термодинамически стресс накапливается в результате действий по раскручиванию ДНК и уплотнению ДНК. Как только гетеродуплекс ДНК-РНК становится достаточно длинным (
10 п.н.), РНК-полимераза освобождает свои вышестоящие контакты и эффективно достигает перехода от промотора к фазе элонгации. Гетеродуплекс в активном центре стабилизирует комплекс элонгации.
ДНК и гены
ДНК ПРОКАРИОТ И ЭУКАРИОТ

Справа крупнейшая спираль ДНК человека, выстроенная из людей на пляже в Варне (Болгария), вошедшая в книгу рекордов Гиннесса 23 апреля 2016 года
Дезоксирибонуклеиновая кислота. Общие сведения
Дезоксирибонуклеи́новая кислота (ДНК) — макромолекула (одна из трёх основных, две другие — РНК и белки), обеспечивающая хранение, передачу из поколения в поколение и реализацию генетической программы развития и функционирования живых организмов. ДНК содержит информацию о структуре различных видов РНК и белков.
В клетках эукариот (животных, растений и грибов) ДНК находится в ядре клетки в составе хромосом, а также в некоторых клеточных органоидах (митохондриях и пластидах). В клетках прокариотических организмов (бактерий и архей) кольцевая или линейная молекула ДНК, так называемый нуклеоид, прикреплена изнутри к клеточной мембране. У них и у низших эукариот (например, дрожжей) встречаются также небольшие автономные, преимущественно кольцевые молекулы ДНК, называемые плазмидами.
С химической точки зрения ДНК — это длинная полимерная молекула, состоящая из повторяющихся блоков — нуклеотидов. Каждый нуклеотид состоит из азотистого основания, сахара (дезоксирибозы) и фосфатной группы. Связи между нуклеотидами в цепи образуются за счёт дезоксирибозы (С) и фосфатной (Ф) группы (фосфодиэфирные связи).

Рис. 2. Нуклертид состоит из азотистого основания, сахара (дезоксирибозы) и фосфатной группы
В подавляющем большинстве случаев (кроме некоторых вирусов, содержащих одноцепочечную ДНК) макромолекула ДНК состоит из двух цепей, ориентированных азотистыми основаниями друг к другу. Эта двухцепочечная молекула закручена по винтовой линии.
В ДНК встречается четыре вида азотистых оснований (аденин, гуанин, тимин и цитозин). Азотистые основания одной из цепей соединены с азотистыми основаниями другой цепи водородными связями согласно принципу комплементарности: аденин соединяется только с тимином (А-Т), гуанин — только с цитозином (Г-Ц). Именно эти пары и составляют «перекладины» винтовой «лестницы» ДНК (см.: рис. 2, 3 и 4).

Рис. 2. Азотистые основания
Последовательность нуклеотидов позволяет «кодировать» информацию о различных типах РНК, наиболее важными из которых являются информационные, или матричные (мРНК), рибосомальные (рРНК) и транспортные (тРНК). Все эти типы РНК синтезируются на матрице ДНК за счёт копирования последовательности ДНК в последовательность РНК, синтезируемой в процессе транскрипции, и принимают участие в биосинтезе белков (процессе трансляции). Помимо кодирующих последовательностей, ДНК клеток содержит последовательности, выполняющие регуляторные и структурные функции.

Рис. 3. Репликация ДНК
Расположение базовых комбинаций химических соединений ДНК и количественные соотношения между этими комбинациями обеспечивают кодирование наследственной информации.
Образование новой ДНК (репликация)
По завершении дупликации образуются две самостоятельные спирали, созданные из химических соединений родительской ДНК и имеющие с ней одинаковый генетический код. Таким путем ДНК способна перерывать информацию от клетки к клетке.
Более подробная информация:
СТРОЕНИЕ НУКЛЕИНОВЫХ КИСЛОТ

Дезоксирибонуклеиновая кислота (ДНК) относится к нуклеиновым кислотам. Нуклеиновые кислоты – это класс нерегулярных биополимеров, мономерами которых являются нуклеотиды.
НУКЛЕОТИДЫ состоят из азотистого основания, соединенного с пятиуглеродным углеводом (пентозой) – дезоксирибозой (в случае ДНК) или рибозой (в случае РНК), который соединяется с остатком фосфорной кислоты (H2PO3–).
Азотистые основания бывают двух типов: пиримидиновые основания – урацил (только в РНК), цитозин и тимин, пуриновые основания – аденин и гуанин.

Рис. 5. Структура нуклеотидов (слева), расположение нуклеотида в ДНК (снизу) и типы азотистых оснований (справа): пиримидиновые и пуриновые

Атомы углерода в молекуле пентозы нумеруются числами от 1 до 5. Фосфат соединяется с третьим и пятым атомами углерода. Так нуклеинотиды соединяются в цепь нуклеиновой кислоты. Таким образом, мы можем выделить 3’ и 5’-концы цепи ДНК:

Рис. 6. Выделение 3’ и 5’-концов цепи ДНК
Две цепи ДНК образуют двойную спираль. Эти цепи в спирали сориентированы в противоположных направлениях. В разных цепях ДНК азотистые основания соединены между собой с помощью водородных связей. Аденин всегда соединяется с тимином, а цитозин – с гуанином. Это называется правилом комплементарности (см. принцип комплементарности ).
Правило комплементарности:
| A–T G–C |
Например, если нам дана цепь ДНК, имеющая последовательность
3’– ATGTCCTAGCTGCTCG – 5’,
то вторая ей цепь будет комплементарна и направлена в противоположном направлении – от 5’-конца к 3’-концу:
5’– TACAGGATCGACGAGC– 3’.

Рис. 7. Направленность цепей молекулы ДНК и соединение азотистых оснований с помощью водородных связей
РЕПЛИКАЦИЯ ДНК
Репликация ДНК – это процесс удвоения молекулы ДНК путем матричного синтеза. В большинстве случаев естественной репликации ДНК праймером для синтеза ДНК является короткий фрагмент РНК (создаваемый заново). Такой рибонуклеотидный праймер создается ферментом праймазой (ДНК-праймаза у прокариот, ДНК-полимераза у эукариот), и впоследствии заменяется дезоксирибонуклеотидами полимеразой, выполняющей в норме функции репарации (исправления химических повреждений и разрывов в молекле ДНК).
Репликация происходит по полуконсервативному механизму. Это значит, что двойная спираль ДНК расплетается и на каждой из ее цепей по принципу комплементарности достраивается новая цепь. Дочерняя молекула ДНК, таким образом, содержит в себе одну цепь от материнской молекулы и одну вновь синтезированную. Репликация происходит в направлении от 3’ к 5’ концу материнской цепи.

Рис. 8. Репликация (удвоение) молекулы ДНК
ДНК-синтез – это не такой сложный процесс, как может показаться на первый взгляд. Если подумать, то для начала нужно разобраться, что же такое синтез. Это процесс объединения чего-либо в одно целое. Образование новой молекулы ДНК проходит в несколько этапов:

Рис. 9. Схематическое изображение процесса репликации ДНК: (1) Отстающая цепь (запаздывающая нить), (2) Ведущая цепь (лидирующая нить), (3) ДНК-полимераза α ( Polα ), (4) ДНК-лигаза, (5) РНК-праймер, (6) Праймаза, (7) Фрагмент Оказаки, (8) ДНК-полимераза δ ( Polδ ), (9) Хеликаза, (10) Однонитевые ДНК-связывающие белки, (11) Топоизомераза.
Далее описан синтез отстающей цепи дочерней ДНК (см. Схему репликативной вилки и функции ферментов репликации)
Нагляднее о репликации ДНК см. видео →
5) Непосредственно сразу после расплетания и стабилизации другой нити материнской молекулы к ней присоединяется ДНК-полимераза α (альфа) и в направлении 5’→3′ синтезирует праймер (РНК-затравку) – последовательность РНК на матрице ДНК длиной от 10 до 200 нуклеотидов. После этого фермент удаляется с нити ДНК.
СТРОЕНИЕ РНК
Рибонуклеиновая кислота (РНК) — одна из трёх основных макромолекул (две другие — ДНК и белки), которые содержатся в клетках всех живых организмов.
Последовательность нуклеотидов позволяет РНК кодировать генетическую информацию. Все клеточные организмы используют РНК (мРНК) для программирования синтеза белков.
Затем матричные РНК (мРНК) принимают участие в процессе, называемом трансляцией, т.е. синтеза белка на матрице мРНК при участии рибосом. Другие РНК после транскрипции подвергаются химическим модификациям, и после образования вторичной и третичной структур выполняют функции, зависящие от типа РНК.

Рис. 10. Отличие ДНК от РНК по азотистому основанию: вместо тимина (Т) в РНК представлен урацил (U), который также комплементарен аденину.
ТРАНСКРИПЦИЯ
Транскрипция – это процесс синтеза РНК на матрице ДНК. ДНК раскручивается на одном из участков. На одной из цепей содержится информация, которую необходимо скопировать на молекулу РНК – эта цепь называется кодирующей. Вторая цепь ДНК, комплементарная кодирующей, называется матричной. В процессе транскрипции на матричной цепи в направлении 3’ – 5’ (по цепи ДНК) синтезируется комплементарная ей цепь РНК. Таким образом, создается РНК-копия кодирующей цепи.
![]()
Рис. 11. Схематическое изображение транскрипции
Например, если нам дана последовательность кодирующей цепи
3’– ATGTCCTAGCTGCTCG – 5’,
то, по правилу комплементарности, матричная цепь будет нести последовательность
5’– TACAGGATCGACGAGC– 3’,
а синтезируемая с нее РНК – последовательность
3’– AUGUCCUAGCUGCUCG – 5’.
ТРАНСЛЯЦИЯ
Рассмотрим механизм синтеза белка на матрице РНК, а также генетический код и его свойства. Также для наглядности по ниже приведенной ссылке рекомендуем посмотреть небольшое видео о процессах транскрипции и трансляции, происходящих в живой клетке:


Рис. 12. Процесс синтеза белка: ДНК кодирует РНК, РНК кодирует белок
ГЕНЕТИЧЕСКИЙ КОД
Генетический код, общий для большинства про- и эукариот. В таблице приведены все 64 кодона и указаны соответствующие аминокислоты. Порядок оснований — от 5′ к 3′ концу мРНК.
Таблица 1. Стандартный генетический код
Среди триплетов есть 4 специальных последовательности, выполняющих функции «знаков препинания»:
Свойства генетического кода
1. Триплетность. Каждая аминокислота кодируется последовательностью из трех нуклеотидов – триплетом или кодоном.

2. Непрерывность. Между триплетами нет никаких дополнительных нуклеотидов, информация считывается непрерывно.

3. Неперекрываемость. Один нуклеотид не может входить одновременно в два триплета.

4. Однозначность. Один кодон может кодировать только одну аминокислоту.

5. Вырожденность. Одна аминокислота может кодироваться несколькими разными кодонами.

6. Универсальность. Генетический код одинаков для всех живых организмов.
Пример. Нам дана последовательность кодирующей цепи:
3’– CCGATTGCACGTCGATCGTATA– 5’.
Матричная цепь будет иметь последовательность:
5’– GGCTAACGTGCAGCTAGCATAT– 3’.
Теперь «синтезируем» с этой цепи информационную РНК:
3’– CCGAUUGCACGUCGAUCGUAUA– 5’.
Синтез белка идет в направлении 5’ → 3’, следовательно, нам нужно перевернуть последовательность, чтобы «прочитать» генетический код:
5’– AUAUGCUAGCUGCACGUUAGCC– 3’.
Теперь найдем старт-кодон AUG:
5’– AU AUG CUAGCUGCACGUUAGCC– 3’.
Разделим последовательность на триплеты:

Найдем стоп-кодон и согласно таблице генетического кода запишем последовательность аминокислот:

Центральная догма молекулярной биологии звучит следующим образом: информация с ДНК передается на РНК (транскрипция), с РНК – на белок (трансляция). ДНК также может удваиваться путем репликации, и также возможен процесс обратной транскрипции, когда по матрице РНК синтезируется ДНК, но такой процесс в основном характерен для вирусов.

Рис. 13. Центральная догма молекулярной биологии
ГЕНОМ: ГЕНЫ и ХРОМОСОМЫ
Термин «геном» был предложен Г. Винклером в 1920 г. для описания совокупности генов, заключенных в гаплоидном наборе хромосом организмов одного биологического вида. Первоначальный смысл этого термина указывал на то, что понятие генома в отличие от генотипа является генетической характеристикой вида в целом, а не отдельной особи. С развитием молекулярной генетики значение данного термина изменилось. Известно, что ДНК, которая является носителем генетической информации у большинства организмов и, следовательно, составляет основу генома, включает в себя не только гены в современном смысле этого слова. Большая часть ДНК эукариотических клеток представлена некодирующими («избыточными») последовательностями нуклеотидов, которые не заключают в себе информации о белках и нуклеиновых кислотах. Таким образом, основную часть генома любого организма составляет вся ДНК его гаплоидного набора хромосом.
Гены — это участки молекул ДНК, кодирующие полипептиды и молекулы РНК
За последнее столетие наше представление о генах существенно изменилось. Ранее геном называли участок хромосомы, кодирующий или определяющий один признак или фенотипическое (видимое) свойство, например цвет глаз.

В 1940 г. Джордж Бидл и Эдвард Тейтем предложили молекулярное определение гена. Ученые обрабатывали споры гриба Neurospora crassa рентгеновским излучением и другими агентами, вызывающими изменения в последовательности ДНК (мутации), и обнаружили мутантные штаммы гриба, утратившие некоторые специфические ферменты, что в некоторых случаях приводило к нарушению целого метаболического пути. Бидл и Тейтем пришли к выводу, что ген — это участок генетического материала, который определяет или кодирует один фермент. Так появилась гипотеза «один ген — один фермент». Позднее эта концепция была расширена до определения «один ген — один полипептид», поскольку многие гены кодируют белки, не являющиеся ферментами, а полипептид может оказаться субъединицей сложного белкового комплекса.
Современное биохимическое определение гена еще более конкретно. Генами называются все участки ДНК, кодирующие первичную последовательность конечных продуктов, к которым относятся полипептиды или РНК, обладающие структурной или каталитической функцией.
Наряду с генами ДНК содержит и другие последовательности, выполняющие исключительно регуляторную функцию. Регуляторные последовательности могут обозначать начало или конец генов, влиять на транскрипцию или указывать место инициации репликации или рекомбинации. Некоторые гены могут экспрессироваться разными путями, при этом один и тот же участок ДНК служит матрицей для образования разных продуктов.
Мы можем приблизительно рассчитать минимальный размер гена, кодирующего средний белок. Каждая аминокислота в полипептидной цепи кодируется последовательностью из трех нуклеотидов; последовательности этих триплетов (кодонов) соответствуют цепочке аминокислот в полипептиде, который кодируется данным геном. Полипептидная цепь из 350 аминокислотных остатков (цепь средней длины) соответствует последовательности из 1050 п.н. (пар нуклеотидов). Однако многие гены эукариот и некоторые гены прокариот прерываются сегментами ДНК, не несущими информации о белке, и поэтому оказываются значительно длиннее, чем показывает простой расчет.
Сколько генов в одной хромосоме?
ДНК прокариот устроена более просто: их клетки не имеют ядра, поэтому ДНК находится непосредственно в цитоплазме в форме нуклеоида.
 Как известно, бактериальные клетки имеют хромосому в виде нити ДНК, уложенной в компактную структуру – нуклеоид. Хромосома прокариота Escherichia coli, чей геном полностью расшифрован, представляет собой кольцевую молекулу ДНК (на самом деле, это не правильный круг, а скорее петля без начала и конца), состоящую из 4 639 675 п.н. В этой последовательности содержится примерно 4300 генов белков и еще 157 генов стабильных молекул РНК. В геноме человека примерно 3,1 млрд пар нуклеотидов, соответствующих почти 29 000 генам, расположенным на 24 разных хромосомах.
Как известно, бактериальные клетки имеют хромосому в виде нити ДНК, уложенной в компактную структуру – нуклеоид. Хромосома прокариота Escherichia coli, чей геном полностью расшифрован, представляет собой кольцевую молекулу ДНК (на самом деле, это не правильный круг, а скорее петля без начала и конца), состоящую из 4 639 675 п.н. В этой последовательности содержится примерно 4300 генов белков и еще 157 генов стабильных молекул РНК. В геноме человека примерно 3,1 млрд пар нуклеотидов, соответствующих почти 29 000 генам, расположенным на 24 разных хромосомах.
Прокариоты (Бактерии).
 Бактерия E. coli имеет одну двухцепочечную кольцевую молекулу ДНК. Она состоит из 4 639 675 п.н. и достигает в длину примерно 1,7 мм, что превышает длину самой клетки E. coli приблизительно в 850 раз. Помимо крупной кольцевой хромосомы в составе нуклеоида многие бактерии содержат одну или несколько маленьких кольцевых молекул ДНК, свободно располагающихся в цитозоле. Такие внехромосомные элементы называют плазмидами (рис. 16).
Бактерия E. coli имеет одну двухцепочечную кольцевую молекулу ДНК. Она состоит из 4 639 675 п.н. и достигает в длину примерно 1,7 мм, что превышает длину самой клетки E. coli приблизительно в 850 раз. Помимо крупной кольцевой хромосомы в составе нуклеоида многие бактерии содержат одну или несколько маленьких кольцевых молекул ДНК, свободно располагающихся в цитозоле. Такие внехромосомные элементы называют плазмидами (рис. 16).
Большинство плазмид состоит всего из нескольких тысяч пар нуклеотидов, некоторые содержат более 10000 п. н. Они несут генетическую информацию и реплицируются с образованием дочерних плазмид, которые попадают в дочерние клетки в процессе деления родительской клетки. Плазмиды обнаружены не только в бактериях, но также в дрожжах и других грибах. Во многих случаях плазмиды не дают никаких преимуществ клеткам-хозяевам, и их единственная задача — независимое воспроизведение. Однако некоторые плазмиды несут полезные для хозяина гены. Например, содержащиеся в плазмидах гены могут придавать клеткам бактерий устойчивость к антибактериальным агентам. Плазмиды, несущие ген β-лактамазы, обеспечивают устойчивость к β-лактамным антибиотикам, таким как пенициллин и амоксициллин. Плазмиды могут переходить от клеток, устойчивых к антибиотикам, к другим клеткам того же или другого вида бактерий, в результате чего эти клетки также становятся резистентными. Интенсивное применение антибиотиков является мощным селективным фактором, способствующим распространению плазмид, кодирующих устойчивость к антибиотикам (а также транспозонов, которые кодируют аналогичные гены) среди болезнетворных бактерий, и приводит к появлению бактериальных штаммов с устойчивостью к нескольким антибиотикам. Врачи начинают понимать опасность широкого использования антибиотиков и назначают их только в случае острой необходимости. По аналогичным причинам ограничивается широкое использование антибиотиков для лечения сельскохозяйственных животных.
Эукариоты.
Таблица 2. ДНК, гены и хромосомы некоторых организмов