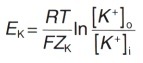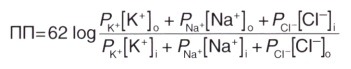Рецепцией называют процесс восприятия и трансформации (преобразования) энергии внешнего раздражителя в энергию нервного импульса или в сложную последовательность внутриклеточных процессов.
Клеточная и сенсорная рецепция
Сенсорной рецепцией называют процесс восприятия и преобразования энергии раздражителей внешней и внутренней среды организма в энергию нервных импульсов, передаваемую по чувствительным нервам в ЦНС. Сенсорный рецептор представляет собой нервную клетку или комплекс нервной и эпителиальной клетки, специально приспособленный для восприятия определенного типа раздражителей. Сенсорные рецепторы являются начальными звеньями любой рефлекторной дуги, а также участвуют в оценке параметров полезного приспособительного результата в функциональных системах организма.
Классификация и строение сенсорных рецепторов
По строению рецепторы подразделяют на первичные и вторичные (рис. 1).
К первичным относят такие сенсорные рецепторы, у которых действие раздражителя воспринимается непосредственно периферическими отростками чувствительного нейрона (нервными окончаниями), которые могут быть:
свободными, т. е. не имеют дополнительных образований;
инкапсулированными, т.е. окончания чувствительного нейрона заключены в особые образования, осуществляющие первичное преобразование энергии раздражителя.
К вторичным относят такие сенсорные рецепторы, у которых действие раздражителя воспринимается специализированной рецептирующей клеткой не нервного происхождения. Возбуждение, возникшее в рецептирующей клетке, передается через синапс на чувствительный нейрон.
Рис. 1. Первичные и вторичные рецепторы.
1 – тело чувствительного нейрона; 2 – периферический отросток чувствительного нейрона (дендрит); 3 – центральный отросток чувствительного нейрона (аксон); 4 – глиальная капсула; 5 – рецептирующая клетка; 6 – синапс между рецептирующей клеткой и чувствительным нейроном.
Тело чувствительного нейрона обычно располагается за пределами ЦНС: в спинномозговом или вегетативном ганглии. От такого нейрона отходят два отростка – дендрит, который следует к периферическим органам и тканям, и аксон, который направляется в спинной мозг.
По расположению сенсорные рецепторы подразделяют на:
экстерорецепторы – воспринимают раздражители из внешней среды организма;
интерорецепторы – воспринимают раздражители из внутренней среды организма;
проприорецепторы – специализированные рецепторы опорнодвигательной системы.
По разнообразию воспринимаемых раздражителей сенсорные рецепторы подразделяют на:
мономодальные – приспособлены для восприятия только одного вида раздражителя;
полимодальные – приспособлены для восприятия различных видов раздражителей.
По модальности сенсорные рецепторы подразделяют на:
хеморецепторы – воспринимают действие химических веществ;
механорецепторы – воспринимают давление, вибрацию, перемещение, степень растяжения;
терморецепторы – чувствительны к изменениям температуры;
ноцицепторы – воспринимают болевое раздражение.
Преобразование энергии в сенсорном рецепторе
Этапы преобразования энергии внешнего раздражителя в энергию нервных импульсов.
Действие раздражителя. Внешний стимул взаимодействует со специфическими мембранными структурами окончаний чувствительного нейрона (в первичном рецепторе) или рецептирующей клетке (во вторичном рецепторе), что приводит к изменению ионной проницаемости мембраны.
Распространение рецепторного потенциала. В первичном рецепторе РП распространяется электротонически и достигает ближайшего перехвата Ранвье. Во вторичном рецепторе РП электротонически распространяется по мембране рецептирующей клетки и достигает пресинаптической мембраны, где вызывает выделение медиатора. В результате срабатывания синапса (между рецептирующей клеткой и чувствительным нейроном) происходит деполяризация постсинаптической мембраны чувствительного нейрона (ВПСП). Образовавшийся ВПСП распространяется электротонически по дендриту чувствительного нейрона и достигает ближайшего перехвата Ранвье.
Закономерности преобразования энергии внешнего раздражителя в серию нервных импульсов (рис. 2):
чем выше сила действующего раздражителя, тем больше амплитуда РП;
чем больше амплитуда РП, тем больше частота нервных импульсов.
Рис. 2. Преобразование энергии внешнего раздражителя в рецепторный потенциал и серию нервных импульсов при действии слабого (А ) и сильного (Б) раздражителя.
Свойства рецепторов
Чувствительность. Количественной мерой чувствительности сенсорного рецептора является абсолютный порог чувствительности – минимальная сила раздражителя, способная вызвать возбуждение рецептора.
Адаптацией называют явление ослабления возбуждения в рецепторе при действии длительного раздражителя постоянной силы.
В зависимости от скорости адаптации рецепторы подразделяют на:
тонические (пропорциональные) рецепторы генерируют нервные импульсы в течение всего времени действия раздражителя; после высокочастотного залпа в начале действия раздражителя частота нервных импульсов устанавливается на постоянном уровне (рис. 3, А);
промежуточные (фазнотонические) рецепторы генерируют нервные импульсы в течение всего времени действия раздражителя, однако их частота существенно уменьшается (рис. 3, Б);
фазные (дифференциальные) рецепторы генерируют нервные импульсы в начальный (ON-ответ) и конечный (OFF-ответ) период действия раздражителя (рис. 4, В).
Рис. 3. Адаптация тонических ( А ), промежуточных ( Б ) и фазных рецепторов ( В ) к длительно действующему раздражителю постоянной силы.
Рецептивное поле
Рецептивным полем нейрона называют множество рецепторов, функционально связанных с этим нейроном. Рецептивное поле нейрона представляет собой динамическое образование – один и тот же нейрон в различные моменты времени может оказаться функционально связанным с различным количеством рецепторов. Максимальная величина рецептивного поля какого-либо нейрона соответствует количеству рецепторов, которые связаны с эти нейроном морфологически, а минимальная величина может ограничиваться всего одним рецептором (рис. 4).
Рис. 4. Рецептивные поля нейронов.
А – максимальное рецептивное поле нейрона 2; Б – минимальное рецептивное поле нейрона 2; В – рецептивное поле нейрона 3; 1 – рецептирующая клетка.
Перекрытие рецептивных полей. У первичных рецепторов зоны ветвления периферических отростков чувствительных нейронов могут перекрывать друг друга (рис. 5, А). У вторичных рецепторов одна рецептирующая клетка может контактировать с несколькими чувствительными нейронами, т. е. может входить в состав рецептивных полей различных нейронов (рис. 6.5, Б).
Плазматическая мембрана всех клеток, в том числе и нейронов, представлена двойным слоем (бислоем) фосфолипидов, в котором «головки» фосфолипидов обращены к водной среде внеклеточных и внутриклеточных пространств, а парные «хвосты» погружены внутрь и образуют липидную мембрану. Фосфатный слой растворим в воде (гидрофильный, или полярный), а двойной липидный слой — нерастворим (гидрофобный, или неполярный).
Внеклеточная и внутриклеточная жидкости представляют собой водно-солевые растворы, в которых множество растворимых молекул диссоциируют на положительно и отрицательно заряженные атомы,— ионы. Ионы и молекулы водных растворов находятся в состоянии постоянного возбуждения и осуществляют процесс диффузии—движения из областей высокой концентрации в область меньшей концентрации. Диффузия ионов происходит не только по градиенту концентрации, но и под влиянием электрических градиентов. Положительно заряженные ионы, например ионы натрия (Na + ) и калия (К + ), называют катионами, поскольку в электрическом поле они движутся к катоду.
• Неуправляемые (независимые) ионные каналы постоянно находятся в открытом состоянии и осуществляют трансмембранный перенос ионов, создавая потенциал покоя на мембранах нейронов.
• Потенциалозависимые (потенциал-управляемые) ионные каналы содержат потенциалчувствительную цепь аминокислот, регулирующую открытие и закрытие отверстий ионных каналов в зависимости от изменений мембранного потенциала. Потенциал-управляемые ионные каналы играют важную роль в формировании потенциала действия.
• Хемоуправляемые (медиаторозависимые) ионные каналы нервной системы обеспечивают временное преобразование мембранного потенциала. В основном эти каналы представлены на постсинаптических мембранах. Активация хемоуправляемых ионных каналов может осуществляться молекулами медиаторов напрямую или опосредованно.
• Механочувствительные каналы активируются под действием физических раздражителей, вызывая деполяризацию мембраны и образование потенциалов действия, что обеспечивает восприятие определенных сигналов нервной системой. Каждый рецептор осуществляет преобразование специфических сигналов, например изменения длины или сократимости мышечных волокон, температурной и тактильной чувствительности кожи, хемочувствительности полости носа и рта или электромагнитных импульсов сетчатки.
На рисунке ниже изображены три неуправляемых ионных канала, обеспечивающих развитие потенциала покоя.
Этот показатель отражает мембранный потенциал покоя, т.е. потенциал мембраны аксона, который не проводит электрические импульсы.
Концентрационный градиент на внешней стороне плазматической мембраны и электрический градиент потенциала на внутренней ее стороне уравновешиваются, когда мембранный потенциал достигает определенной величины, которую называют равновесным потенциалом для ионов К + (Ek). Его величина может быть рассчитана из уравнения Нернста, которое позволяет представить электрический градиент потенциала иона через его концентрационный градиент на основании законов термодинамики:
где Ek — равновесный потенциал для ионов К + (мВ)
R — газовая постоянная (8,31 Дж/(моль/°К)
Т — абсолютная температура в градусах Кельвина (310 К = 37 °С) F — число Фарадея (96500 Кл/моль)
Zk — валентность ионов К + (+1)
ln — натуральный логарифм
[К + ]0 — концентрация ионов K + с наружной стороны мембраны
[К + ]i — концентрация ионов К + с внутренней стороны мембраны.
Переведем натуральный логарифм в десятичный и рассчитаем значение показателя дроби:
Для расчета мембранного потенциала покоя применяют уравнение Гольдмана, учитывающее проницаемость мембраны для трех основных ионов и их приблизительное количество.
где ПП — потенциал покоя
62 — RT/Fx2,3 (постоянная для перевода натурального логарифма в десятичный)
2. Натрий-калиевый насос. Под влиянием градиента концентрации происходит непрерывное перемещение ионов Na + внутрь клетки, а ионов К + — наружу, что нарушает постоянство потенциала покоя. Поддержание этого равновесия обеспечивает натрий-калиевый насос, корректирующий пассивное движение ионов. Натрий-калиевый насос представляет собой канал, способный перемещать ионы Na + наружу, а ионы К + — внутрь клетки. Во время работы насоса происходит перенос трех ионов Na + из клетки на каждые два иона К + в клетку.
Перенос обоих катионов осуществляется против градиента концентрации за счет энергии превращения АТФ в АДФ ферментом АТФазой. Активация этого фермента происходит при повышении концентрации ионов Na + в цитозоле.
Редактор: Искандер Милевски. Дата публикации: 11.11.2018
Клеточные механизмы восприятия вкуса
Об авторе
Роман Александрович Романов — доктор биологических наук, старший научный сотрудник лаборатории молекулярной физиологии клетки Института биофизики клетки РАН (Пущино, Московская обл.). Научные интересы связаны с механизмами афферентной нейропередачи во вкусовых клетках.
Рис. 1. Вкусовая система [1]
Строение периферического отдела вкусовой системы
Рис. 2. Схематическое изображение вкусовой почки [1]
Этот отдел включает более 5000 особых образований — сосочков разной формы: грибовидных (они находятся в передней части языка), желобовидных (в его корневой доли) и листовидных (по бокам). Уже на этом уровне проявляется определенная, хотя и неполная, специализация. Так, основная информация о сладком приходит в мозг прежде всего от грибовидных сосочков, тогда как о горьком — от желобовидных, а о кислом и соленом — от листовидных. Вкусовые сосочки состоят из почек (в общей сложности их во рту около 10 тыс.) — ассоциатов из 50–100 выстроенных подобно долькам апельсина специализированных клеток вытянутой формы (рис. 2). Они обладают многими свойствами нейронов, такими как способность к генерации потенциалов действия (волн возбуждения, возникающих, как правило, при деполяризации мембраны, изменении ее потенциала в сторону положительных значений) и образование синапсов (мест контакта между двумя клетками). Но, несмотря на наличие признаков электровозбудимых тканей, вкусовые, как и сенсорные клетки органов слуха и равновесия, развиваются из эпителия. Сама вкусовая почка не достигает поверхности слизистой оболочки языка, в полость рта выходит только пора, через которую растворенные в слюне вещества контактируют со вкусовыми клетками. Их наружная часть представлена микроворсинками с расположенными на них рецепторами — главными участниками процессов распознавания, усиления и преобразования пищевых сигналов. Рецепторы микроворсинок при связывании своих лигандов (веществ, специфически соединяющихся с рецептором) запускают каскад биохимических реакций во вкусовой клетке, которая в ответ высвобождает нейромедиатор (химическое вещество, передающее электрический сигнал на нервное волокно), стимулирующий вкусовой нерв. И в итоге в мозг уходят электрические импульсы, несущие информацию об интенсивности и разновидности воздействия.
Среди вкусовых клеток выделяют как минимум три типа: темные (I), так как они интенсивно окрашиваются на ультратонких срезах, светлые (II) и промежуточные (III). Апикальная часть, непосредственно контактирующая с ротовой полостью, обычно богата микроворсинками (исключение составляют клетки III типа, у них только одна микроворсинка) и несет метаботропные рецепторы (вызывающие изменения в клеточном метаболизме после связывания со своими лигандами), сопряженные с G-белками, ионные каналы и другие сигнальные белки. Базальная часть, не имеющая доступа ко вкусовым веществам, отвечает за связь с афферентными и эфферентными (проводящими возбуждение от органов и тканей в центральную нервную систему и наоборот) нервными окончаниями и, возможно, с другими рецепторными клетками вкусовой почки. Апикальные части всех трех типов клеток соединены друг с другом окружающими их плотными контактами — белковыми образованиями, которые выглядят как пояс из переплетающихся нитей и служат барьером для диффузии вкусовых веществ. Средняя продолжительность жизни вкусовой клетки не превышает 10 дней.
Функциональные типы вкусовых клеток
В 2000-х годах во вкусовых клетках удалось идентифицировать ряд важных белков, а главное, определить их роль в восприятии стимулов той или иной модальности. Были открыты и молекулярные рецепторы горьких и сладких веществ, принадлежащие разным семействам. Сейчас известно около 30 членов семейства T2R (Taste Receptors), специфичных для вкусовых клеток метаботропных G-белковых рецепторов, которые чувствительны к горькому [2]. Их области, находящиеся вне клетки (это потенциальные сайты связывания с лигандами), т. е. экспонированные в ротовую полость, сильно различаются аминокислотной последовательностью, а остальная часть этих молекул высококонсервативна. В одной клетке у млекопитающих одновременно работает множество T2R-рецепторов для различных горьких веществ, действующих на одни и те же элементы внутриклеточного сигнального каскада. Вот почему среди горького не существует различающихся «подвкусий», все горькое ощущается одинаково. Совсем иначе воспринимается сладкое — углеводы, белки и аминокислоты, синтетические соединения. Все они возбуждают один-единственный рецептор, который обладает очень большой внеклеточной частью с множеством аллостерических центров, связывающих сладкие вещества [3]. Этот универсальный для всего спектра сладкого рецептор формируется двумя белками из другого специфического для вкусовых клеток семейства — T1R, в результате образуется димер T1R2+T1R3.
С помощью электронной микроскопии, иммуногистохимии и РНК-гибридизации in situ в популяции вкусовых клеток того или иного типа постепенно были идентифицированы различные белки, участвующие в кодировании (переводе интенсивности и длительности раздражения в электрический импульс) вкусовых стимулов. Благодаря этому удалось выявить физиологические функции клеток и понять, что их типы отличаются не только морфологически, но и функционально (рис. 3). Выяснилось, что клетки II типа подразделяются на самом деле на три отдельные группы, ответственные за восприятие горького (T2R), сладкого (димеры T1R2+T1R3) и умами (димеры T1R1+T1R3). Однако для всех трех групп общей остается система внутриклеточной сигнализации, запускаемая в ответ на вкусовую стимуляцию. Каскад реакций включает активацию G-белка гастдуцина и фосфолипазы С β2, увеличение концентрации кальция в цитозоле и открытие кальций-зависимого ионного канала TRPM5, влекущее за собой деполяризацию мембраны [4]. Вкусовые клетки III типа ответственны за восприятие кислого, поскольку именно в них функционирует pH-активируемый ионный канал PKD2L1. Это удалось доказать в экспериментах на генетически модифицированных мышах — они теряли чувствительность к кислому в отсутствие клеток, экспрессирующих PKD2L1 [5]. Клетки I типа, с одной стороны, выполняют функции глиальных (создающих специфическое микроокружение для нейронов) и отчасти сходны с ними морфологически — имеют отростки, правда, крыловидные. Последние создают физические барьеры между клетками вкусовой почки и предотвращают вытекание нейромедиатора из пространства с нервными окончаниями. Более того, на плазматической мембране клеток I типа есть характерный для глии белок — транспортер глутаминовой и аспарагиновой аминокислот (GLAST — GLutamate ASpartate Transporter), а также внеклеточный фермент эктоапираза 2, или эктонуклеозидтрифосфатдифосфогидролаза (E-NTPDase — EctoNucleoside TriPhosphate Diphosphohydrolase) [6, 7]. Они утилизируют молекулы глутамата и АТФ, появляющиеся в межклеточном пространстве вкусовой почки в процессе акта нейропередачи. Считается, что АТФ высвобождается вкусовыми клетками при их стимуляции, а глутамат — эфферентными нервными окончаниями, которые регулируют работу вкусовой почки. С другой стороны, клетки I типа участвуют в рецепции соленого, так как именно в них функционируют эпителиальные натриевые каналы (ENaC — Epithelial Na + Сhannel), блокируемые диуретиком амилоридом, а соленое восприятие им в значительной степени регулируется.
Рис. 3. Морфофункциональные типы клеток вкусовой почки. Клетки I типа выполняют функции глиальных, так как они имеют аминокислотный транспортер GLAST и эктоапиразу NTPDase-2, и участвуют в рецепции соленого за счет натриевых каналов ENaC. Клетки II типа подразделяются на три группы, ответственные за восприятие горького (благодаря семейству рецепторов T2R), сладкого (участвуют димеры T1R2+T1R3) и умами (обеспечивается димерами T1R1+T1R3). Клетками III типа воспринимается кислое за счет pH-активируемых ионных каналов PKD2L1
Афферентная нейропередача во вкусовых клетках
Как же вкусовые клетки разных типов передают информацию на нервные окончания? Обычно нейропередача происходит благодаря синаптическим пузырькам, наполненным афферентным нейромедиатором. При слиянии с клеточной мембраной (кальций-зависимом экзоцитозе) они высвобождают свое содержимое в межклеточное пространство, возбуждая близлежащее нервное окончание. За открытие молекулярных основ везикулярного транспорта Джеймсу Ротману, Ренди Шекману и Томасу Зюдофу в 2013 г. была присуждена Нобелевская премия по физиологии или медицине. Однако только в клетках III типа, распознающих кислое, имеются синаптические пузырьки и другие атрибуты классических химических синапсов, включая белки комплекса SNARE (они осуществляют экзоцитоз) и потенциал-зависимые кальциевые каналы (благодаря которым процесс экзоцитоза запускается). Ранее, кстати, из-за того что образование синапсов с афферентным нервным окончанием присуще лишь клеткам III типа, они ошибочно рассматривались как единственные истинно рецепторные. Кислые вещества вызывают открытие ионных каналов PKD2L1 и других протон-чувствительных каналов клеток III типа. Входящий ток (обусловленный поступлением в клетку ионов) приводит к деполяризации мембраны выше критического уровня и к генерации серии потенциалов действия, частота которых зависит от силы тока, пропорционального, в свою очередь, концентрации протонов во вкусовой поре. Это влечет за собой активацию потенциал-зависимых кальциевых каналов и вход кальция в клетку. Повышение его концентрации в цитозоле запускает экзоцитоз синаптических пузырьков и высвобождение нейромедиатора серотонина на вкусовое нервное окончание. Таким образом, нейропередача клетками III типа осуществляется вполне традиционным способом (рис. 4).
Рис. 4. Нейропередача вкусовыми клетками III типа. Кислые вещества вызывают открытие ионных каналов PKD2L1 и других протон-чувствительных каналов клеток III типа. Входящий ток (обусловленный поступлением в клетку ионов) приводит к деполяризации мембраны выше критического уровня и генерации серии потенциалов действия, частота которых зависит от силы тока, пропорционального, в свою очередь, концентрации протонов во вкусовой поре. Это влечет за собой активацию потенциал-зависимых (ПЗ) кальциевых каналов и вход кальция в клетку. Повышение его концентрации в цитозоле запускает экзоцитоз синаптических пузырьков и высвобождение нейромедиатора серотонина на вкусовое нервное окончание
Сложнее и запутаннее обстоит дело с другими клетками. Синаптических контактов с нервными волокнами клетки I типа не формируют. Считается, что их основная функция во вкусовой почке — поддерживающая, вспомогательная. Поэтому механизм, используемый ими для передачи в мозг информации о соленом, до сих пор не исследован. До недавнего времени также совершенно неизученными оставались механизмы афферентной нейропередачи во вкусовых клетках II типа. Хотя это основные рецепторные клетки для горького, сладкого и умами, в них отсутствуют синаптические структуры (включая белки комплекса SNARE и потенциал-зависимые кальциевые каналы). Еще одна проблема заключается в том, что клетки II типа электрически возбудимы, но, поскольку они не образуют классических синапсов, совершенно непонятно, для чего ими генерируются потенциалы действия. Афферентным нейромедиатором, передающим информацию о вкусе от этих клеток, оказался АТФ. Уже более 20 лет известно, что АТФ — это не только макроэргическая молекула, универсальный источник энергии для различных биохимических и физиологических процессов, но и внеклеточный лиганд множества рецепторов [8]. В качестве нейромедиатора АТФ, например, участвует в болевой чувствительности (многие знают не понаслышке, что подкожные инъекции АТФ весьма болезненны). В 2005 г. группа исследователей установила, что афферентные нейроны мышей, у которых были одновременно инактивированы гены, кодирующие АТФ-рецепторы P2X2 и P2X3, перестают отвечать на вкусовые стимулы всех модальностей, а реакции таких животных на сладкое, горькое и умами сильно снижены [9]. Это свидетельствовало о роли АТФ в качестве вкусового нейромедиатора и подтверждало наблюдение, что горькие вещества вызывают секрецию АТФ из языкового эпителия. Возникло предположение, что цепь событий, приводящих к выбросу нейромедиатора и возбуждению вкусового нерва в ответ на горькое, сладкое и умами, должна включать — как необходимые этапы — активацию ионного канала TRPM5 (согласно данным по генетическому нокауту и свойствам рекомбинантного, т. е. полученного с помощью генной инженерии, канала) и клеточную деполяризацию. Значит, выброс клеткой нейромедиатора, АТФ, должен стимулироваться деполяризацией мембраны. Связь ответа на горькое и деполяризации уже наблюдали ранее в экспериментах со стимуляцией вкусовых клеток смесью двух горьких веществ — циклогексимида и денатония.
Мы проверили способность вкусовых клеток, выделенных из языка мыши, высвобождать АТФ в ответ на деполяризацию мембраны. В экспериментах мы одновременно управляли электрической активностью клеток и измеряли концентрацию кальция в АТФ-биосенсорах. В качестве последних использовались клетки линии COS-1, загруженные кальциевым индикатором Fluo-4. Они проявляют уникально высокую чувствительность к АТФ в среде, генерируя внутриклеточные кальциевые ответы (т. е. флуоресцируя под микроскопом) при концентрациях АТФ от 50 нМ, и крайне низкую чувствительность к ряду других нейромедиаторов, ранее идентифицированных во вкусовой почке. Типы вкусовых клеток мы различали по их электрофизиологическим свойствам (форме ионных токов, генерируемых в ответ на ряд последовательных, ступенчатых поляризаций от −100 до 50 мВ), которые регистрировались непосредственно в ходе эксперимента [10–12]. Вкусовые клетки II типа при потенциале покоя (мембранном потенциале невозбужденной клетки, т. е. разности электрических потенциалов на внутренней и наружной сторонах мембраны) в диапазоне −70. −40 мВ не вызывали в клетках линии COS-1, расположенных рядом, никаких кальциевых ответов. Но деполяризация клеток II типа до потенциала −10 мВ и выше вызывала повышение концентрации кальция в клетках COS-1 с задержкой от 3 до 10 с — в зависимости от расстояния между вкусовой клеткой и АТФ-биосенсором (рис. 5). Подавление флуоресценции АТФ-биосенсора в присутствии ингибитора АТФ-рецепторов сурамина подтвердило, что нейромедиатором, высвобождаемым вкусовыми клетками II типа, служит именно АТФ [11].
Рис. 5. Схема эксперимента, подтверждающего высвобождение АТФ вкусовыми клетками II типа (а–г). Вкусовой клеткой при потенциале −70 мВ АТФ не секретируется, и потому расположенные рядом с ней АТФ-биосенсоры (клетки линии COS-1) не флуоресцируют (б). Однако после деполяризации вкусовой клетки до 10 мВ в АТФ-биосенсорах увеличивается концентрация кальция, что видно по усилению в них флуоресценции кальциевого зонда Fluo-4 (в). Восстановление исходного потенциала вкусовой клетки (−70 мВ) приводит к падению флуоресценции АТФ-биосенсоров (г). Цветовая шкала (в, справа) показывает интенсивность флуоресценции, увеличивающуюся от синего к красному. Адаптировано из [11]
Секреция АТФ может осуществляться двумя известными на сегодня способами: либо везикулярно (т. е. посредством экзоцитоза, управляемого локальным повышением внутриклеточной концентрации кальция), либо через специфические мембранные транспортеры или ионные каналы [13]. Предположим, что во вкусовых клетках II типа работает везикулярный механизм выброса АТФ. Тогда, если заблокировать вход внешнего кальция в клетку и/или не дать ей самой увеличить концентрацию кальция в цитозоле, секреция АТФ должна заметно снизиться. Но уменьшение концентрации кальция в среде до 100 нМ и введение в клетки II типа быстрого кальциевого хелатора BAPTA (связывающего ионы кальция) не привели к заметным изменениям в секреции ими АТФ. Значит, его высвобождение происходит не классическим везикулярным (кальций-зависимым) способом, а при участии ионных каналов [11]. Если это действительно так, то они должны быть проницаемы не только для АТФ, молекулярная масса которого около 600 Да, но и для других, более мелких молекул, например, флуоресцентных трейсеров (веществ, используемых для наблюдения за протеканием химической реакции или биологического процесса). Так, каналы, формируемые белками коннексинами, проницаемы для молекул массой до 1 кДа, включая АТФ, цАМФ (циклический аденозинмонофосфат), инозитолтрифосфат (IP3 — Inositol trisPhosphate) и др. Поэтому гипотезу высвобождения АТФ через ионные каналы можно подтвердить, доказав, что в ответ на деполяризацию внутрь вкусовых клеток проникают крупные отрицательно заряженные флуоресцентные молекулы из окружающего раствора. И действительно, в наших экспериментах красители Lucifer Yellow (LY), флуоресцеин и карбоксифлуоресцеин оказывались внутри вкусовых клеток II типа после их деполяризации — они начинали ярко светиться под флуоресцентным микроскопом [11, 14]. Итак, выброс АТФ и вход в клетки II типа флуоресцентных красителей зависят от потенциала. Деполяризация необходима для адекватного вкусового восприятия, но сама по себе не приводит к увеличению концентрации кальция в цитозоле (в отличие от клеток III типа с потенциал-зависимыми кальциевыми каналами). Все это свидетельствует в пользу того, что АТФ во вкусовых клетках II типа высвобождается через специфические потенциал-зависимые ионные каналы.
Интересно было разобраться, какие белки формируют во вкусовых клетках II типа столь необычные АТФ-проницаемые ионные каналы. На момент исследований кандидатами служили некоторые анионные каналы, а также полупоры, образованные белками коннексинами и паннексином 1. Известные блокаторы анионных каналов и ингибиторы паннексина 1 не оказывали значительных эффектов на секрецию АТФ [11]. Она сохранялась и у животных с выключенным геном, кодирующим паннексин 1, в ответ на вкусовую и электрическую стимуляцию [11, 14, 15]. Это исключало возможность участия анионных каналов и паннексина 1 в высвобождении АТФ клетками II типа. Далее мы проверили влияние на их АТФ-проницаемые каналы ряда веществ, которые ингибируют открытие различных полупор. Только октанол, неспецифичный ингибитор коннексинов, и пептид 43 GAP26, также блокирующий некоторые коннексины, сильно подавляли выброс АТФ. Похоже было, что роль АТФ-транспортера во вкусовых клетках выполняет канал, образованный коннексинами. Окончательную ясность в этот вопрос внесла группа зарубежных ученых. Они доказали, что АТФ на самом деле транспортируется недавно открытым потенциал-зависимым ионным каналом CALHM1 (CALcium Homeostasis Modulator 1), который просто похож на коннексины некоторыми свойствами [16]. Этот канал, присутствующий только в клетках II типа, оказался проницаемым для АТФ. Мыши с генетическим нокаутом гена, кодирующего CALHM1, теряли чувствительность к горькому, сладкому и умами [17]. Во вкусовых клетках II типа таких мышей сильно уменьшались потенциал-зависимые токи, а секреция АТФ вкусовыми почками снижалась.
Необычный механизм передачи информации на нервное окончание, реализуемый во вкусовых клетках II типа (в отсутствие цепочки «потенциал-зависимая активация кальциевых каналов — экзоцитоз синаптических пузырьков — высвобождение нейромедиатора»), заставил усомниться в роли потенциалов действия в афферентной нейропередаче. Обычно вторичные сенсорные клетки (непосредственно воспринимающие раздражение и затем передающие сигнал о нем на нервную клетку), не имеющие аксонов, не используют потенциалы действия для передачи информации и даже не способны их генерировать. Но вкусовые клетки II типа млекопитающих определенно генерируют потенциалы действия в ответ и на электрическое, и на пищевое воздействие. Зачем? Не исключено, что у потенциалов действия существует еще одна физиологическая функция — регуляция невезикулярного высвобождения нейромедиатора. Важное свойство секреции АТФ клетками этого типа — ее сильная зависимость от мембранного потенциала. Поэтому логично было предположить, что потенциалы действия управляют афферентной нейропередачей. Экспериментально мы установили, что при характерном для клеток II типа потенциале покоя от −55 до −40 мВ деполяризация до −45. −30 мВ, индуцируемая активацией канала TRPM5, не вызывает высвобождение АТФ. Но когда она сопровождается генерацией потенциалов действия, частота или количество которых пропорциональны ее величине, уровень секретированного ATФ оказывается пропорциональным интенсивности вкусового сигнала. Это обеспечивает адекватное кодирование сенсорной информации. За счет резкой зависимости количества ATФ от потенциала фактически происходит квантовый (т. е. унифицированный по кинетике и величине) выброс ATФ через неселективные ионные каналы в ответ на каждый потенциал действия.
Мы дополнительно доказали участие потенциалов действия в высвобождении ATФ клетками II типа и другим способом, — вызывая его кратковременными деполяризационными импульсами, сопоставимыми по длительности с потенциалами действия [11]. По результатам работ японских авторов, блокатор натриевых каналов тетродотоксин вызывает заметное снижение уровня АТФ, выбрасываемого в ответ на стимуляцию вкусовых клеток горькими веществами [18]. Количество АТФ при этом пропорционально числу сгенерированных потенциалов действия.
Рис. 6. Гипотетическая последовательность событий при рецепции вкусовых веществ клетками II типа. Вкусовые молекулы связываются с рецепторами, находящимися в апикальной части клетки. Это последовательно приводит к активации G-белков, фосфолипазы С β2 (PLC β2) и к мобилизации внутриклеточного кальция, благодаря которому открывается кальций-зависимый ионный канал TRPM5, что вызывает осцилляционную деполяризацию мембраны и генерацию серии потенциалов действия. Это стимулирует потенциал-зависимые (ПЗ) ионные каналы CALHM1, проницаемые для АТФ, и ведет к его высвобождению. Он активирует рецепторы P2X афферентных нервных окончаний, а генерация серии потенциалов действия позволяет закодировать информацию об интенсивности вкусового воздействия. Вкусовая клетка I типа гидролизует АТФ, высвобождаемый на нервное окончание, с помощью эктоапиразы NTPD-ase2. Кроме того, внешний АТФ приводит к мобилизации внутриклеточного кальция в клетке I типа при участии метаботропных P2Y-рецепторов и фосфоинозитидного каскада, включающего фосфолипазу С (PLC), инозитолтрифосфат (IP3) и диацилглицерол (DAG). Стимулируется также и кальций-активируемый анионный канал
Многие, несомненно, задаются вопросом: зачем изучать механизмы вкусового восприятия? Действительно ли нам необходимо знать, какие клетки и как в нем участвуют? Оказывается, значительное число людей страдает расстройством вкуса. У большинства оно развивается после травмы головы или инфекции верхних дыхательных путей и среднего уха; после лучевой терапии (при лечении онкологических заболеваний головы и шеи); в результате действия определенных химикатов (инсектицидов и некоторых лекарств, включая антибиотики и антигистаминные средства) и как последствие некоторых видов хирургии уха, носа и горла. Опасны ли расстройства вкуса? Да, так как они ослабляют или вовсе устраняют врожденную систему предупреждения об испорченной пище, а иногда и об аллергенах. При ухудшении чувства вкуса мы можем начать есть слишком мало / много и терять / набирать вес. Потеря вкуса может вынудить нас употреблять чрезмерные количества сахара или соли, чтобы улучшить вкус пищи, а для людей с сахарным диабетом или высоким кровяным давлением это очень рискованно. В тяжелых случаях потеря вкуса может даже привести к депрессии. Поэтому знания о механизмах его восприятия необходимы для изучения причин, приводящих к отклонениям, и поиска способов их коррекции. А это — важная научная и медицинская задача.
Литература 1. Благутина В. В. Анатомия вкуса // Химия и жизнь. 2010. № 10. C. 34–38. 2. Lindemann B. Receptors and transduction in taste // Nature. 2001. V. 413. P. 219–225. 3. Margolskee R. F. Molecular mechanisms of bitter and sweet taste transduction // J. Biol. Chem. 2002. V. 277. P. 1–4. 4. Zhang Y., Hoon M. A., Chandrashekar J. et al. Coding of sweet, bitter, and umami tastes, different receptor cells sharing similar signaling pathways // Cell. 2003. V. 112. P. 293–301. 5. Huang A. L., Chen X., Hoon M. A. et al. The cells and logic for mammalian sour taste detection // Nature. 2006. V. 442. P. 934–938. 6. Bartel D. L., Sullivan S. L., Lavoie E. G. et al. Nucleoside triphosphate diphosphohydrolase-2 is the Ecto-ATPase of type I cells in taste buds // J. Comp. Neurol. 2006. V. 497. P. 1–12. 7. Lawton D. M., Furness D. N., Lindemann B. et al. Localization of the glutamate-aspartate transporter, GLAST, in rat taste buds // Eur. J. Neurosci. 2000. V. 12. P. 3163–3671. 8. Ralevic V., Burnstock G. Receptors for purines and pyrimidines // Pharmacol. Rev. 1998. V. 50. P. 413–492. 9. Finger T. E., Danilova V., Barrows J. et al. ATP signaling is crucial for communication from taste buds to gustatory nerves // Science. 2005. V. 310. P. 1495–1499. 10. Romanov R. A., Kolesnikov S. S. Electrophysiologicaly identified subpopulations of taste bud cells // Neurosci. Lett. 2006. V. 395. P. 249–254. 11. Romanov R. A., Rogachevskaja O. A., Bystrova M. F. et al. Afferent neurotransmission mediated by hemichannels in mammalian taste cells // EMBO J. 2007. V. 26. P. 657–667. 12. Bystrova M. F., Romanov R. A., Rogachevskaya O. A. et al. Functional expression of the extracellular calcium-sensing receptor in mouse taste cells // J. Cell Sci. 2010. V. 123. P. 972–982. 13. Lazarowski E. R., Boucher R. C., Harden T. K. Mechanisms of release of nucleotides and integration of their action as P2X- and P2Y-receptor activating molecules // Mol. Pharmacol. 2003. V. 64. P. 785–795. 14. Romanov R. A., Bystrova M. F., Rogachevskaya O. A. et al. The ATP permeability of pannexin 1 channels in a heterologous system and in mammalian taste cells is dispensable // J. Cell Sci. 2012. V. 125. P. 5514–5523. 15. Romanov R. A., Rogachevskaja O. A., Khokhlov A. A. et al. Voltage-dependence of ATP secretion in mammalian taste cells // J. Gen. Physiol. 2008. V. 132. P. 731–744. 16. Siebert A. P., Ma Z., Grevet J. D. et al. Structural and functional similarities of calcium homeostasis modulator 1 (CALHM1) ion channel with connexins, pannexins, and innexins // J. Biol. Chem. 2013. V. 288. P. 6140–6153. 17. Taruno A., Vingtdeux V., Ohmoto M. et al. CALHM1 ion channel mediates purinergic neurotransmission of sweet, bitter and umami tastes // Nature. 2013. V. 495. P. 223–226. 18. Murata Y., Yasuo T., Yoshida R. et al. Action potential-enhanced ATP release from taste cells through hemichannels // J. Neurophysiol. 2010. V. 104. P. 896–901.





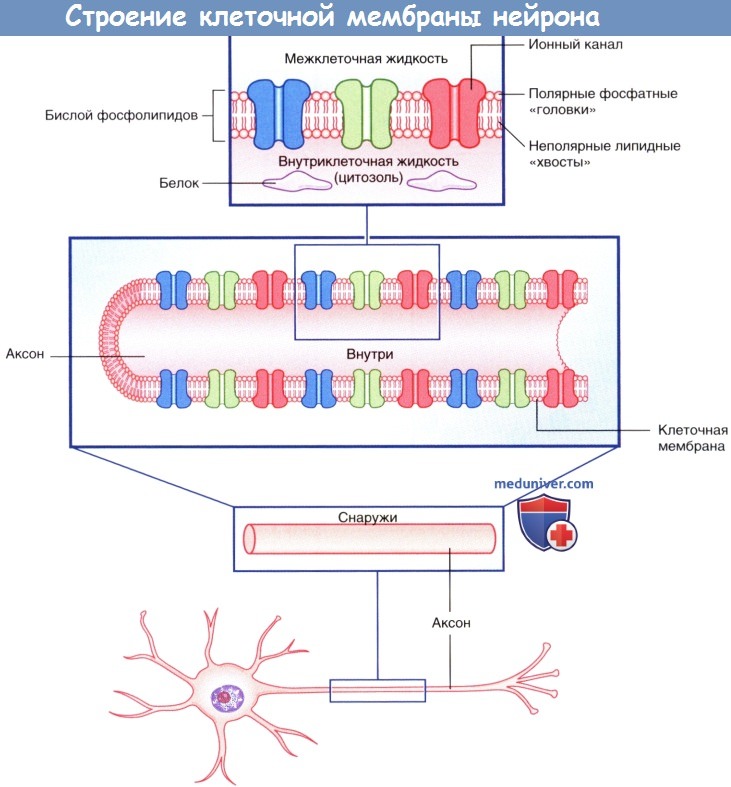 Строение клеточной мембраны нейрона.
Строение клеточной мембраны нейрона.